
Hematose

Após a ventilação dos alvéolos ocorre a difusão de oxigênio dos alvéolos para o sangue pulmonar e a difusão do dióxido de carbono na direção oposta. A difusão ocorre devido ao movimento cinético das moléculas dos gases. A velocidade de difusão de cada um dos gases participantes da respiração é diretamente proporcional à pressão causada por este gás, chamada de pressão parcial do gás.
Cada gás contribui para a pressão total em proporção direta à sua concentração. Os gases dissolvidos na água e nos tecidos do corpo também exercem pressão, porque as moléculas dissolvidas estão em movimento aleatório e têm energia cinética. Quando o ar penetra nas vias respiratórias, a água das superfícies dessas vias imediatamente se evapora e umedece o ar. Isto é resultado do fato de que as moléculas de água, como as diferentes moléculas de gases dissolvidos, estão continuamente escapando da superfície de água para a fase gasosa.
A pressão que as moléculas de água exercem para escapar através da superfície é chamada de pressão de vapor da água. A difusão efetiva de um gás de área de alta pressão para área de baixa pressão é igual ao número de moléculas que se movimentam nesta direção menos o número que se movimenta na direção oposta, e isto, por sua vez, é proporcional à diferença entre a pressão de gás das duas áreas, chamada de diferença de pressão de difusão.
Todos os gases que têm importância respiratória são altamente solúveis em lipídios e, conseqüentemente, altamente solúveis nas membranas celulares. O ar alveolar não tem a mesma concentração de gases que o ar atmosférico, devido ao fato de que a cada ciclo respiratório o ar alveolar é parcialmente renovado pelo ar atmosférico, o oxigênio está constantemente sendo absorvido do ar alveolar e o dióxido de carbono se difundindo do sangue pulmonar para os alvéolos.
À medida que entra nas vias respiratórias, o ar é exposto aos líquidos que revestem as superfícies respiratórias e é totalmente umidificado antes de entrar nos alvéolos. Somente 350 ml de ar fresco são trazidos para os alvéolos em cada inspiração normal e a mesma quantidade é eliminada a cada expiração, de modo que muitas inspirações são necessárias para substituir a maior parte do ar alveolar. Esta substituição lenta do ar alveolar é importante para impedir mudanças bruscas nas concentrações gasosas do sangue.
O oxigênio está constantemente sendo absorvido pelo sangue dos pulmões, e oxigênio novo, da atmosfera, está continuamente sendo inspirado pelos alvéolos. Quanto mais rapidamente o oxigênio é absorvido, mais baixa será sua concentração nos alvéolos; por outro lado, quanto mais rápido o oxigênio novo é inspirado pelos alvéolos, mais alta será sua concentração. O dióxido de carbono é continuamente formado no organismo, em seguida descarregado nos alvéolos e removido pela ventilação.
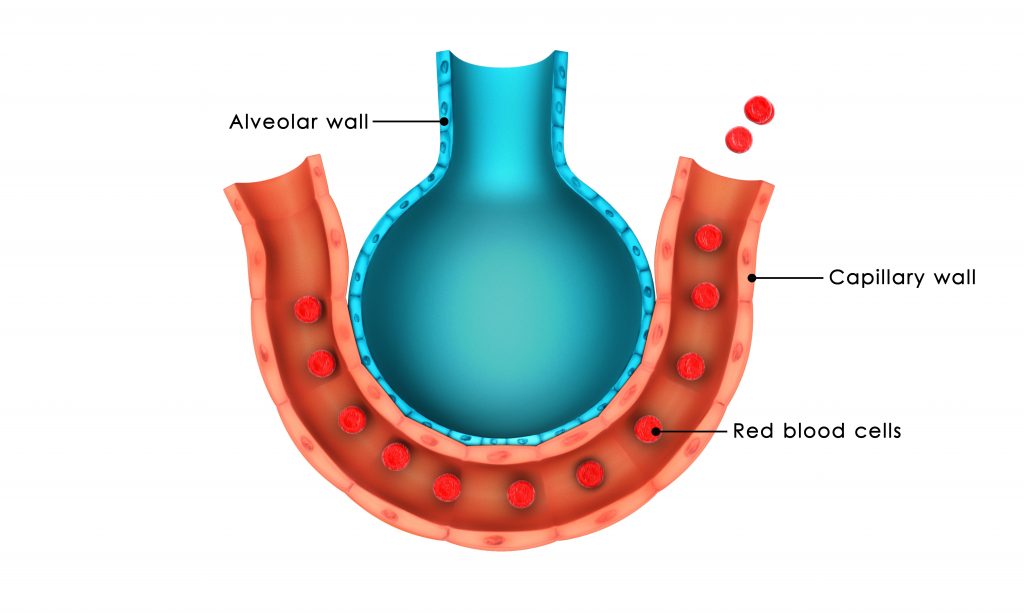
As concentrações e pressões de oxigênio e de dióxido de carbono nos alvéolos são determinadas pelas velocidades de absorção ou de excreção dos dois gases e também pelo nível de ventilação alveolar. A unidade respiratória é formada por um bronquíolo respiratório, ductos alveolares, átrios e alvéolos. As paredes destas estruturas possuem uma extensa rede de capilares interconectados, conhecida como membrana respiratória.
A partir de estudos histológicos estima-se que a superfície total da membrana respiratória tenha área de aproximadamente 50 a 100 metros quadrados no adulto normal. Os principais fatores que a velocidade da difusão gasosa através da membrana respiratória são a espessura da membrana, a área superficial da membrana, a velocidade de difusão do gás e a diferença de pressão entre os dois lados da membrana.
Quando o sangue arterial alcança os tecidos periféricos, sua pressão parcial de oxigênio é maior do que a pressão parcial de oxigênio no líquido intersticial. Essa enorme diferença de pressão causa a difusão muito rápida do oxigênio do sangue para os tecidos. Quando o oxigênio é utilizado pelas células, a maior parte dele é transformada em dióxido de carbono e este aumenta a sua pressão parcial intracelular.
Em seguida, o dióxido de carbono se difunde das células para os capilares teciduais e depois é levado pelo sangue para os pulmões, onde se difunde dos capilares pulmonares para os alvéolos. Normalmente, cerca de 97% do oxigênio transportado dos pulmões para os tecidos é carregado em combinação química com a hemoglobina nas hemácias, e os 3% restantes são transportados dissolvidos na água do plasma e das células.
Assim, em condições normais, o oxigênio é transportado para os tecidos quase totalmente pela hemoglobina. Quando a pressão parcial de oxigênio está alta, como nos capilares pulmonares, o oxigênio de liga com a hemoglobina, mas quando a pressão parcial de oxigênio está baixa, como nos capilares teciduais, o oxigênio é liberado da hemoglobina. Esta é a base para quase todo o transporte de oxigênio dos pulmões para os tecidos.
Sob condições normais, a velocidade de utilização de oxigênio pelas células é controlada, em última análise, pela velocidade de consumo energético dentro das células, isto é, pela velocidade com que o ADP é produzido a partir do ATP. O monóxido de carbono se combina com a hemoglobina no mesmo ponto onde o oxigênio se associa e, por conseguinte, pode deslocar o oxigênio da hemoglobina.
Além disso, ele se liga à hemoglobina com 250 vezes mais firmeza que o oxigênio. Um paciente gravemente envenenado com monóxido de carbono pode ser adequadamente tratado administrando-se oxigênio puro, pois o oxigênio em altas pressões alveolares desloca o monóxido de carbono mais rapidamente do que o oxigênio sob baixa pressão atmosférica.
O dióxido de carbono pode ser transportado sob a forma dissolvida (7%), combinando-se com a água no interior das hemácias para formar ácido carbônico e, em seguida os íons hidrogênio e bicarbonato catalizado pela anidrase carbônica (70%) e combinado com a hemoglobina e proteínas plasmáticas (15 a 25%). O ácido carbônico formado quando o dióxido de carbono entra no sangue dos tecidos diminui o pH sanguíneo.
Contudo, a reação deste ácido com os tampões do sangue impede que a concentração de íons hidrogênio aumente muito (e que o pH desça muito). Normalmente, o sangue arterial tem um pH de aproximadamente 7,41 e, à medida que o sangue adquire dióxido de carbono nos capilares teciduais, o pH desce para um valor de aproximadamente 7,37. Ocorre o reverso quando o dióxido de carbono é liberado do sangue para os pulmões, com o pH se elevando para o valor arterial.
Regulação da Respiração
O centro respiratório é composto de vários grupos de neurônios localizados bilateralmente no bulbo e na ponte. É dividido em três grandes grupos de neurônios: (1) um grupo dorsal respiratório, localizado na região dorsal do bulbo, responsável principalmente pela inspiração, (2) um grupo ventral respiratório, localizado na região ventrolateral do bulbo, responsável tanto pela expiração quanto pela inspiração, dependendo dos neurônios que são estimulados e (3) o centro pneumotáxico, localizado dorsalmente na região superior da ponte, e que ajuda a controlar tanto a freqüência quanto o padrão da respiração.
O grupo dorsal respiratório de neurônios desempenha um papel fundamental no controle da respiração. Ele se estende ao longo da maior parte do comprimento do bulbo. Todos ou quase todos os seus neurônios estão localizados no núcleo do tracto solitário, embora neurônios adicionais da substância reticular adjacente ao bulbo provavelmente também desempenhem papéis importantes no controle respiratório.
O núcleo do tracto solitário também é uma terminação sensorial dos nervos vago e glossofaríngeo, que transmitem sinais sensoriais dos quimioceptores, dos baroceptores e de vários tipos diferentes de receptores pulmonares para o centro respiratório. O ritmo básico da respiração é gerado principalmente no grupo dorsal respiratório de neurônios. Mesmo quando todas as terminações nervosas periféricas que entram no bulbo são seccionadas e o tronco encefálico também é seccionado acima e abaixo do bulbo, este grupo de neurônios ainda emite, repetitivamente, potenciais de ação inspiratórios.
A causa básica dessas descargas repetitivas, porém, ainda é desconhecida. O sinal inspiratório ocorre “em rampa”, iniciando-se muito fraco e aumentando progressivamente por cerca de dois segundos. Em seguida, cessa abruptamente por cerca de três segundos e permite a retração elástica da caixa torácica e dos pulmões causando a expiração. O centro pneumotáxico limita a duração da inspiração e aumenta a freqüência respiratória.
O grupo ventral de neurônios permanece quase totalmente inativo durante a respiração normal em repouso. Quando há necessidade de altos níveis de ventilação pulmonar, essa área opera mais ou menos como um mecanismo multiplicador. Dessa forma, o grupo ventral é essencialmente importante na respiração forçada. Os receptores de estiramento localizados nas paredes dos brônquios e bronquíolos, que transmitem os sinais através dos nervos vagos para o grupo respiratório dorsal quando os pulmões ficam muito distendidos ativam uma resposta de feedback adequada que desliga a rampa inspiratória através do chamado reflexo de insuflação de Hering-Breuer.
O último objetivo da respiração é manter as concentrações adequadas de oxigênio, dióxido de carbono e íons hidrogênio nos tecidos. Portanto, é importante que a atividade respiratória seja altamente responsiva às variações de cada um desses elementos. O excesso de dióxido de carbono ou de íons hidrogênio causa aumento na intensidade dos sinais inspiratórios e expiratórios para os músculos da respiração. O oxigênio atua quase totalmente nos quimioceptores periféricos localizados nos corpos carotídeos e aórticos, e estes transmitem sinais adequados para o centro respiratório através do nervo de Hering.
A área quimiossensitiva do centro respiratório é muito sensível às mudanças da pressão parcial de dióxido de carbono na circulação ou à concentração de íons hidrogênio. Os neurônios da área sensitiva são principalmente sensíveis aos íons hidrogênio. Entretanto, este íon tem dificuldade em atravessar as barreiras hemoencefálica e hemoliquórica, ao contrário do dióxido de carbono. Embora o dióxido de carbono tenha pouco efeito direto na estimulação dos neurônios da área quimiossensitiva, ele exerce um potente efeito indireto. Este efeito é resultado da reação do dióxido de carbono com a água dos tecidos para formar ácido carbônico.
Este, por sua vez, se dissocia em íons hidrogênio e íons bicarbonato; os íons hidrogênio têm potente efeito estimulador direto. Quando uma pessoa respira ar com muito pouco oxigênio, isto, obviamente, diminui a pressão parcial de oxigênio sanguínea e excita os quimioceptores carotídeos e aórticos, desse modo aumentando a respiração. Entretanto, esse efeito é muito menor do que se espera, porque o aumento da respiração remove o dióxido de carbono dos pulmões e conseqüentemente diminui a pressão parcial de dióxido de carbono e a concentração de íons hidrogênio do sangue.
Estas duas alterações deprimem intensamente o centro respiratório, como foi discutido anteriormente, de modo que o efeito final dos quimioceptores em aumentar a respiração em resposta à diminuição da pressão parcial de oxigênio é totalmente contrabalançado. A causa mais predominante de depressão respiratória e parada respiratória seja talvez a dosagem excessiva de anestésicos e narcóticos.
O pentabarbital sódico e a morfina deprimem intensamente o centro respiratório. Outros anestésicos como o halotano são mais comumente utilizados por não apresentarem estas características de forma tão acentuada. Uma anormalidade da respiração chamada de respiração periódica ocorre em diferentes enfermidades.
A pessoa respira profundamente por um pequeno intervalo de tempo e depois respira fracamente ou não respira durante um intervalo, e este ciclo se repete continuadamente. O tipo mais comum de respiração periódica é a respiração de Cheyne-Stokes, caracterizada por aumento e diminuição lenta da respiração que se repetem aproximadamente a cada 40 a 60 segundos.
